


Community Ecology and the Science of Coexistence
Reflections on Academic Ecology and What We Still Have to Learn

This article contains the thoughts I have been developing over many years regarding the value and limitations of modern approaches to community ecology. Through a reflective essay and some poetic musings, I have aimed to share some of the deeper truths I have come to understand during my time studying the ecology of plant communities. Whether you choose to read my poetry, my prose, or both, I hope these words help you appreciate the beauty, vitality, and importance of our complex, diverse, and shared community of life
Reflections on Academic Plant Community Ecology
Over the past 5 years, I have been struggling to understand how ecosystems change. I have been seeking an answer: Why do we see these plants here? And further, why do these plants seem to love living together? After I discovered my keen interest in the beauty and diversity of plant life, I was fortunate enough to be welcomed into an academic research lab working to understand plant community ecology. Through my time in this lab — which is now coming to a close — I dove into the rich intricacy of community ecology. I became surrounded by the various scholars, numerous species and study systems, and new conceptual frameworks through which I could come to answer these questions. Since I began in this lab, I have grappled with the technical frameworks and quantitative tools used to explore plant ecological diversity. Through this process, I have come to realize some of the inherent limitations in these scientific tools and ways of knowing.
Within this lab, we study how and why various species of plants coexist together on the landscape. To explore these, we work within two key paradigms: functional trait ecology and modern coexistence theory. Through these frameworks, we work to understand which parts of the plants we study cause them to use different resources and occupy distinct ecological niches – which we can then use to explain why plant species can coexist.
To apply functional trait ecology to explore plant diversity and coexistence, we first need to identify discrete characteristics (whether they be morphological, developmental, chemical, or otherwise) that can be measured on a diverse suite of plants. The variation observed in these traits are then analyzed to understand the axes of ecological variation they describe. By analyzing functional traits — often including plant height, rooting depth, root and crown architecture, phenology (ie. when a plant first starts growing, when it blooms, and when it sets seed), among others — we can understand ecological differences within and between species, allowing us to understand how plants acclimate and adapt to the environment in which they live.
Generally, the traits we rely on are widely found within plants, able to be easily quantified, and map clearly onto the functional ability of a plant to survive and reproduce in their environment. This paradigm has its merits – namely, we can assess variation within and between species, and we can analyze how these differences coincide with environmental variation and niche in which an organism lives. However, not all functionally relevant characteristics are easily quantified or widely comparable. For example, in working to understand how invasive annual grasses compare to the native plants, qualitative variation in the photosynthetic pathway, in response to disturbance, in phenology, and in growth form (ie. grass vs. forb vs. shrub vs. tree) leaves large gaps in our ability to understand key ecological differences between species.
Beyond functional traits, I have also been struggling to understand the mathematical models, assumptions, and implications of modern coexistence theory: a framework for coexistence between two species. This model is built on the assumption of competitive exclusions – that two species cannot coexist unless they occupy distinct enough niches (if you would like to dig into this framework, feel free to check out this open-source review article). Modern coexistence theory aims to show how differences in the niche or fitness of two species can allow for the two species to coexist. Unless a species, when it becomes rare, acquires some competitive advantage (such as not competing with individuals, including other individuals of the same species, which rely on the exact same set and proportions of resources), a species will not be able to coexist with the others in the community.
This framework for understanding coexistence has some glaring limitations. First, this framework only allows us to determine whether two species are able to coexist. This would be fine, except reality is full of complex ecosystems in which many species co-occur. Some of the communities with the greatest diversity are also the productive and resilient to disturbance and change.
The second limitation is that through this framework, we reduce coexistence to a quantitative mathematical equation. While we still rely on key empirical measurements — derived from experiments in which the fecundity of two species is assessed at varying densities of the other — we lose sight of the bigger picture of community-scale coexistence. This approach allows us to understand how they respond to the competitive pressure imposed by one another, but doesn’t allow us to explore how these might change if the climate or the environment was different. With the sheer magnitude of species known to coexist, and higher-order (effects beyond those predictable with pairwise models) interactions known to stabilize or disrupt coexistence, this approach lacks explanatory power. We remain unable to predict real diversity of coexisting plant species in the communities we observe across the natural world.
The third limitation of this framework is its reliance on common garden experiments (all species being grown in the same area, experiencing the same soil, climate, and biotic environment). This requirement limits our ability to understand the complex interactions between species, many of which facilitate coexistence. For example, if symbiotic microbial species live in the soil, and differentially benefit the nutrient availability or root health of one or both species, our expectation for coexistence or competitive exclusion can fail to accord with real observed interactions. Symbiotic relationships — long-term interactions sustained between two or more species where at least one comes to rely on the interspecific (between species) relationship — can help ensure a plant has its needs met without directly competing with its neighbors for resources. Without acknowledging these relationships, it becomes very difficult to understand how, for instance, common shared fungal networks or other shared symbiotic relationships help transform neighborhood interactions, shifting relationships that would otherwise be competitive into collaborative, mutualistic ones.
The fundamental limitations of these paradigms all stem from the overarching mission of “Western” science: to reduce complex phenomena into their constituent parts. And yet, the whole is often not merely the sum of its parts. By aspiring to reduce the tremendous qualitative diversity observed in the natural world into mathematically tractable models, we can lose track of the deeper reality of coexistence. Coexistence can become something we seek to prove, not something we work to understand as intrinsic to healthy communities.
We thus fail to understand the truth of community ecology: all life is entangled with the other living and non-living environment that makes up their neighborhood; all life shapes its neighborhood, generally by negotiating beneficial (or at least minimally harmful) relationships with their neighbors; all life is interdependent with other beings. To understand the niche occupied by any organism, we must understand the symbiotic relationships which collectively shape its environment.
From a Narrow Scope to Broad Notions of the Ecological Community
Many community ecologists only ever study their particular group of organisms. Some study the mammal community – or even just the Carnivoran (mammals belonging to the order Carnivora) community. Others study bird communities, insect communities, or microbial communities – whatever cohesive and distinct collection of organisms can easily understood to interact, compete or collaborate, and coexist with one another. Yet, in studying a community of organisms, we know that symbioses — key relationships between species — bring organisms from distant branches on the tree of life into collaborative (and sometimes parasitic) relations. It is impossible to understand why a plant community is the way it is without integrating it with the broader living community: from bacteria and fungi, to animals of all sorts, plants depend on other forms of life at every moment of their lives. It is impossible to study the community of an ecosystem without working to understand how all relate to all.
As a plant community ecologist, my focus begins at the “bottom” of the ecosystem — where energy and nutrients enter the trophic “pyramid” or the food web — the network of interactions, which move energy and nutrients throughout the ecosystem. I found my passion for plants because through appreciating their diversity — both their beauty and their diverse places on the landscape — that I have come to understand the diversity of habitats that make up our home. Through understanding the ecology of various plant species, I have come to see how different landscapes’ soil and water, hills and valleys, human and non-human life, sets the stage for particular plants to thrive. Together, a community of plants can teach us about the land – they can tell us where water is scarce or where water rises near the surface, they can remind us of the land’s history, human land-uses, and environment change; and they can teach us what it might take to become truly at home here, wherever we might be.
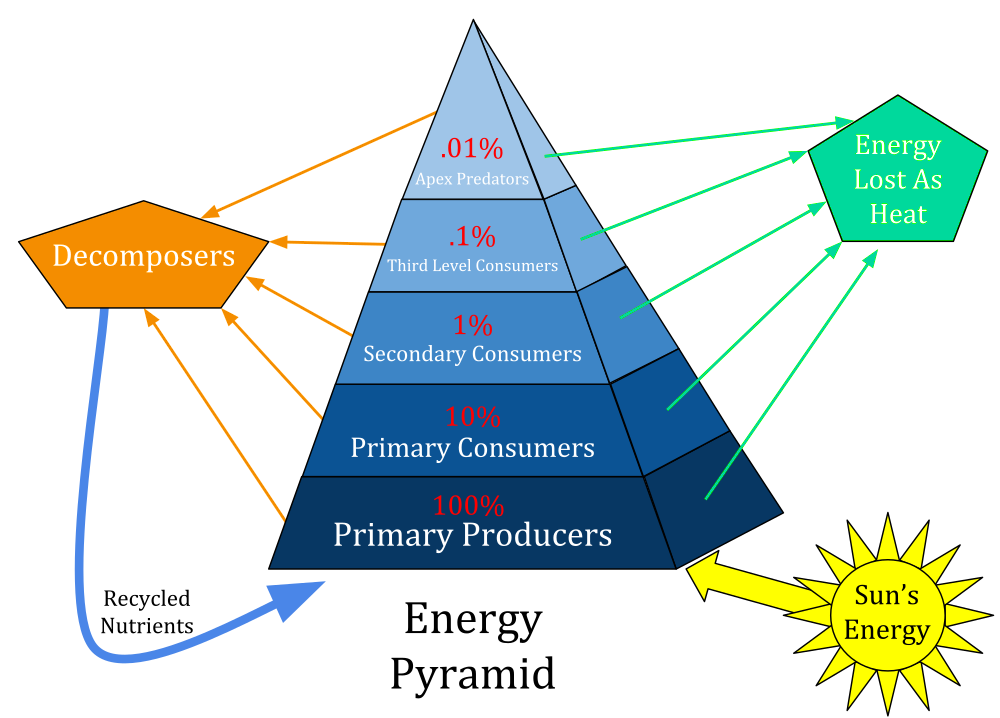
This diagram shows the loss of biomass as we move up the “trophic levels” – only about 10% is retained as we move up the food chain. While I have been trained as a plant community ecologist, my Master’s research centered the interactions which link together the plant, bacterial, and fungal community – bringing me into contact with the foundational tie between soil nutrients, primary producers, and decomposers. But without considering the long history of human belonging and engagement in the ecosystem, the current state of the ecosystem could not be understood.
Now, after 5 years of learning, I realize that the approach we use to understand plant communities and coexistence is profoundly limited. We cannot understand how and why plants live together without understanding the broader history and context – weaving together the complex story of coevolution between plants and animals, soils and microbes each collaborating and competing to establish their place in their shared home; this requires that we not only reckon with inherent differences between organisms, but that we also recognize the profound impact that human land-uses and power structures have had on this land. We cannot understand a plant community solely by dissecting each plant species, each pairwise interaction, into a set of quantitative values: there are things that go beyond what is easily measured and compared. To understand the ecology and evolution of a community, we must grapple with the whole.
Towards an Integrative Approach to Community Ecology
I first recognized these limitations when I realized that every variable we assessed was quantitative – and yet the process of niche construction, including the various ways in which an organism might shape its environment to better suit itself and its offspring, forces us to explore qualitative steps in evolution. A more holistic approach perspective on the relationships between organisms, their neighborhood, and the wider environment requires we integrate niche construction with coexistence
I grew up in the Redwood forest; a landscape where the trees tower overhead, shading the forest floor from which Redwood seedlings sprout and grow. In the rich leaf litter shed by their ancestors (and in many cases, other trunks belonging to the same or a closely related tree), and the moist, shaded environment of the understory, Redwood saplings are ideally positioned to get established and thrive. High above, the crowns of the surrounding Redwoods reach into the sky, leaves outstretched under the cloudy sky, where sunlight and water ensure carbon can be incorporated into the forest below. These trees actively shape the climate of this environment – seeding and pulling in fog, ensuring the habitat remains ideal for forest’s trees.
The Redwoods guide this process: they cannot be easily broken into parts, quantified, and compared with other trees in the environment. And why should it need to?

Gazing up at crowns of Redwood trees will always inspire a sense of awe and wonder. These ancient trees always remind me of the deep intelligence, rootedness, and interconnectedness of all life, sheltering the forest floor under a canopy of loving warmth.
The community that makes up the life of an ecosystem is composed of innumerable relationships within and between different species, each with its own way of life, each with its own set of needs and goals – serving innumerable functions and providing uncountable resources for others to make use of.
Resources and functions – these are the words we used to describe how an organism supports the wider ecosystem. The soil provides resources and serves a number of functions (which some call “ecological services”), and in turn, each organism does the same. There is no ecological community without an interwoven network of resources and functions – at each step along a food web, carbon becomes fixed as part of an organism’s body, only to again become the food that others will eat. Organic matter: a constant flux from life to death to life renewed.
From the living churn of nutrients, incorporated and made available by bacterial metabolism and the branching webs of fungi in the soil, and throughout the diverse lifeways of insects, birds, mammals, and other animals, each organism in the ecological community is wholly interdependent on the broader ecosystem – and from one perspective, it all comes back to the plants. There is no plant community without the broader living community.
Within Jewish Ecology, we must reckon with the unity of Life – there is one community, one World; all life is both a part of this larger whole, and yet each organism is a whole in themselves. No life can survive alone. Only in dynamic relations with neighbors and our wider home can life of any kind survive and thrive.
Ultimately, all human life is anchored within the living community. No socio-cultural group is detached from this reality – and neither is any species. Just as no person can live apart from other human beings, no life can live isolated from the living world. Community ecology, as an academic discipline, may today remain disparate and divided, broken up into various sub disciplines – specializations on various taxonomic groups, ecosystems, and functional groups. And yet, the realization that the human community is just a part of the broader more-than-human community continues to percolate through science; it is still just taking root in academic ecology. May this wisdom bloom in our lifetime.
My time learning community ecology has inspired a deep appreciation for the real significance of interdependence; of coexistence. Still, I remain struck by the lack of substantive integration regarding these ideas. If we are to learn to coexist with other ways of being and knowing, other cultures and other species, we must recognize that coexistence is not something we need to prove: it is foundational to life as a whole. We observe the world, across social and natural history, throughout landscapes today, and we see immense diversity. Researchers acknowledge this: coexistence and competitive exclusion are two assumptions from which community ecology research can stem, and today we have a choice – Is diversity or exclusion the natural state of our community?
And so, we — as ecologically concerned citizens of the world — have a responsibility to answer this question, even if it means just picking an assumption we prefer. Do different species and cultures necessarily lead to competition, exclusion, and elimination? Or is there a natural tendency to diverge from our competitors, to work towards complementary relationships; to evolve persistent diversity and coexist? Perhaps, Life as a whole — from the earliest single-celled organisms to the multi-trillion celled animals and fungi which have diversified and spread across our living world — has a tendency towards diversification, differentiation, and integration. And even if it also has a tendency to move towards domination, uniformity, and division — as the only species capable of imagining otherwise, do we not have a responsibility to work towards unity, integration, and coexistence? Only together, in working towards mutual understanding, substantive solidarity, and genuine care, can we work towards a world in which all can live together in peace and in harmony. Perhaps it's time for our science to understand this.
A Poem on Coexistence
The following poem seeks to convey some of the deeper insights I have come to understand regarding what allows our living community to survive, thrive, and sustainably evolve. I hope these words resonate with you.
Living Together
Life drives difference,
Difference drives multi-functionality;
Integration.
Integration drives Unity,
But Unity can breed uniformity.
Uniformity leads to assimilation;
Assimilation and narrow-minded economics
Breeds destruction,
Colonization,
Of body and mind,
Spirit and Earth.
Destruction.
It leads us to hate people who trespass our imagined boundaries of
Gender,
Race,
Nation.
Borders.
Imagined categories rendered real.
Separation
Destroys our humanity.
We can we fail to let others find their own way.
We work against the freedom of life,
We limit our capacity and willingness to love.
We fail each other.
We fail ourself.
We lose our ability to engage life as a whole,
We make ourselves less resilient,
We pillage the Earth.
We endanger the future of trillions of lives now,
But most of all,
We lose sight of our highest calling;
Our fundamental responsibility
To all life:
To live and love,
To carry forth the knowledge and memories of our forebears;
our understandings of History and the Science of Nature;
To work for the sake of the future.
We must learn to coexist,
Cocreate,
To coevolve in complementarity.
Diversity is power.
Power in People,
Power in all Nature,
Together we are resilient;
We can learn to work for All.
For peace and harmony,
For all our ancestors,
And for all life yet to live.
Living together,
Here,
For the sake of All.
One community
One home
We have no other home.
Humanity,
Latin name: Homo sapiens
Translation: the great, wise ape
Our humanity is grounded in our capacity to learn from others,
To work together,
With Life,
Land,
Water,
And Seed;
To love
To liberate,
And to bring forth
A sustainable and mutualistic relationship with all Life.
We must realize that intelligence can take on many voices,
Many faces,
Many forms.
Collective intelligence,
Co-creation,
The greatest wisdom,
The highest form of human knowledge,
Is rooted in the many millions of years of collaborative story-making.
Ecology and History may help us understand what our capacity is.
But we must listen to,
And learn from,
The many stories of the World.
Truth can only be found if we listen to other’s stories.
Other Peoples,
Other lineages,
Branches of this Great Tree of Life.
There is deep knowledge and wisdom,
And intelligence in the collaborative relationships between people,
Plants,
Fungi,
and other ecosystems.
Domestication: to bring a species into the domus (latin, the home).
We have made the Earth our home.
Humanity has become at home,
Implicated in nearly every environment,
Every habitat,
All the Earth.
We must work to love our home.
Every body,
Every ecosystem,
Every species,
To love the whole process of life;
that which has brought forth and uncountable number cells, species, and beings,
And continues in Us.
In all of Nature.
Our Destiny is to truly know that our love for our neighbors,
And our love for G-d are one and the same.
To take responsibility for our power,
To work to make our home a place,
Where all other beings,
Other lives,
Can belong.
The health of all life –
for an uncountable many generations to come
depends on Us.
We must love all lives,
All Nature;
And find ways to work
In make free
Healthy,
And Whole
The reality of every person,
Every People,
Every loving relation,
In every Land,
Every River,
And Every Sea
Here on the Earth.
Through Science and Philosophy, compassion and love, we find ourselves fundamentally here, together, living for the sake of all life. Whether in poetry or prose, art or reason, politics or religion, we have the capacity to engage with and to express our care for our community. Through reading these reflections, I hope that you have a deeper appreciation for the resilience, productivity, and fundamental interdependence that lies within the diversity of our ecological community. Through these words, I hope to have moved beyond the detached objectivity of science, and through the inherent subjectivity of poetry – I hope to have struck a chord that continues to reverberate for the days to come.
Thank you for reading Jewish Ecology. This blog aims to build a participatory dialogue in which important ideas at the intersection of Judaism and ecology can help us root ourselves in the World. This blog is not just for Jews; I aim for these articles to be accessible and impactful for anyone that is interested in building up an understanding of the role that spirituality, philosophy, and/or ethics play in bringing about a more peaceful, ecological, and sustainable future for all People and the World.
Thank you for the poem; it's not only beautiful and powerful, but it shares so much love!